生物化學與分子生物學/糖酵解途徑
| 醫學電子書 >> 《生物化學與分子生物學》 >> 糖代謝 >> 糖的分解代謝 >> 糖酵解途徑 |
| 生物化學與分子生物學 |
|
|
糖酵解途徑是指細胞在胞漿中分解葡萄糖生成丙酮酸(pyruvate)的過程,此過程中伴有少量ATP的生成。在缺氧條件下丙酮酸被還原為乳酸(lactate)稱為糖酵解。有氧條件下丙酮酸可進一步氧化分解生成乙酰CoA進入三羧酸循環,生成CO2和H2O。
目錄 |
(一)葡萄糖的轉運(transport of glucose)
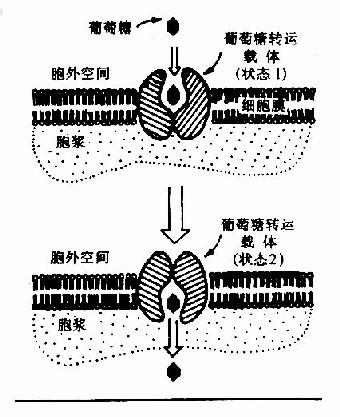
圖4-1 葡萄糖通過轉運載體轉入細胞示意圖 GLUT代表葡萄糖轉運載體
葡萄糖不能直接擴散進入細胞內,其通過兩種方式轉運入細胞:一種是在前一節提到的與Na+共轉運方式,它是一個耗能逆濃度梯度轉運,主要發生在小腸粘膜細胞、腎小管上皮細胞等部位;另一種方式是通過細胞膜上特定轉運載體將葡萄糖轉運入細胞內(圖4-1),它是一個不耗能順濃度梯度的轉運過程。目前已知轉運載體有5種,其具有組織特異性如轉運載體-1(GLUT-1)主要存在于紅細胞,而轉運載體-4(GLUT-4)主要存在于脂肪組織和肌肉組織。
(二)糖酵解過程
糖酵解分為兩個階段共10個反應,每個分子葡萄糖經第一階段共5個反應,消耗2個分子ATP為耗能過程,第二階段5個反應生成4個分子ATP為釋能過程。
1.第一階段
(1)葡萄糖的磷酸化(phosphorylation of glucose)
進入細胞內的葡萄糖首先在第6位碳上被磷酸化生成6-磷酸葡萄糖(glucose6phophate,G-6-P),磷酸根由ATP供給,這一過程不僅活化了葡萄糖,有利于它進一步參與合成與分解代謝,同時還能使進入細胞的葡萄糖不再逸出細胞。催化此反應的酶是己糖激酶(hexokinase,HK)。己糖激酶催化的反應不可逆,反應需要消耗能量ATP,Mg2+是反應的激活劑,它能催化葡萄糖、甘露糖、氨基葡萄糖、果糖進行不可逆的磷酸化反應,生成相應的6-磷酸酯,6-磷酸葡萄糖是HK的反饋抑制物,此酶是糖氧化反應過程的限速酶(ratelimiting enzyme)或稱關鍵酶(key enzyme)它有同工酶Ⅰ-Ⅳ型,Ⅰ、Ⅱ、Ⅲ型主要存在于肝外組織,其對葡萄糖Km值為10-5~10-6M。
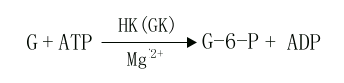
Ⅳ型主要存在于肝臟,特稱葡萄糖激酶(glucokinase,GK),對葡萄糖的Km值1~10-2M,正常血糖濃度為5mmol/L,當血糖濃度升高時,GK活性增加,葡萄糖和胰島素能誘導肝臟合成GK,GK能催化葡萄糖、甘露糖生成其6-磷酸酯,6-磷酸葡萄糖對此酶無抑制作用。
HK與GK兩者區別見表4-1。
表4-1 己糖激酶(HK)和葡萄糖激酶(GK)的區別
| HK | GK | |
| 組織分布 | 絕大多數組織 | 肝臟和β細胞 |
| Km | 低 | 高 |
| 6-磷酸葡萄糖的抑制 | 有 | 無 |
(2)6-磷酸葡萄糖的異構反應(isomerization of glucose-6-phosphate)
這是由磷酸己糖異構酶(phosphohexoseisomerase)催化6-磷酸葡萄糖(醛糖aldose sugar)轉變為6-磷酸果糖(fructose-6-phosphate,F-6-P)的過程,此反應是可逆的。

(3)6-磷酸果糖的磷酸化(phosphorylation of fructose-6-phosphate)
此反應是6磷酸果糖第一位上的C進一步磷酸化生成1,6-二磷酸果糖,磷酸根由ATP供給,催化此反應的酶是磷酸果糖激酶1(phosphofructokinasel,PFK1)。
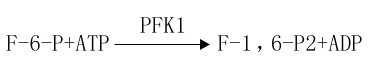
PFK1催化的反應是不可逆反應,它是糖的有氧氧化過程中最重要的限速酶,它也是變構酶,檸檬酸、ATP等是變構抑制劑,ADP、AMP、Pi、1,6-二磷酸果糖等是變構激活劑,胰島素可誘導它的生成。
(4)1.6二磷酸果糖裂解反應(cleavageof fructose1,6 di/bis phosphate)
醛縮酶(aldolase)催化1.6-二磷酸果糖生成磷酸二羥丙酮和3-磷酸甘油醛,此反應是可逆的。

(5)磷酸二羥丙酮的異構反應(isomerization of dihydroxyacetonephosphate)
磷酸丙糖異構酶(triosephosphate isomerase)催化磷酸二羥丙酮轉變為3-磷酸甘油醛,此反應也是可逆的。
![]()
到此1分子葡萄糖生成2分子3-磷酸甘油醛,通過兩次磷酸化作用消耗2分子ATP。
2.第二階段:
(6)3-磷酸甘油醛氧化反應(oxidation of glyceraldehyde-3-phosphate
此反應由3-磷酸甘油醛脫氫酶(glyceraldehyde 3-phosphatedehydrogenase)催化3-磷酸甘油醛氧化脫氫并磷酸化生成含有1個高能磷酸鍵的1,3-二磷酸甘油酸,本反應脫下的氫和電子轉給脫氫酶的輔酶NAD+生成NADH+H+,磷酸根來自無機磷酸。

(7)1.3-二磷酸甘油酸的高能磷酸鍵轉移反應
在磷酸甘油酸激酶(phosphaglyceratekinase,PGK)催化下,1.3-二磷酸甘油酸生成3-磷酸甘油酸,同時其C1上的高能磷酸根轉移給ADP生成ATP,這種底物氧化過程中產生的能量直接將ADP磷酸化生成ATP的過程,稱為底物水平磷酸化(substratelevel phosphorylation)。此激酶催化的反應是可逆的。
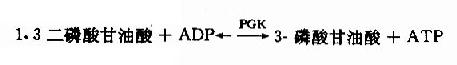
(8)3-磷酸甘油酸的變位反應
在磷酸甘油酸變位酶(phosphoglyceratemutase)催化下3-磷酸甘油酸C3-位上的磷酸基轉變到C2位上生成2-磷酸甘油酸。此反應是可逆的。

(9)2-磷酸甘油酸的脫水反應
由烯醇化酶(enolase)催化,2-磷酸甘油酸脫水的同時,能量重新分配,生成含高能磷酸鍵的磷酸烯醇式丙酮酸(phosphoenolpyruvatePEP)。本反應也是可逆的。
![]()
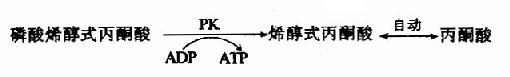
(10)磷酸烯醇式丙酮酸的磷酸轉移
在丙酮酸激酶(pyruvatekinase,PK)催化下,磷酸烯醇式丙酮酸上的高能磷酸根轉移至ADP生成ATP,這是又一次底物水平上的磷酸化過程。但此反應是不可逆的。
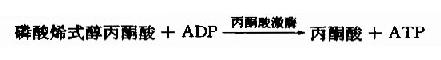
丙酮酸激酶是糖的有氧氧化過程中的限速酶,具有變構酶性質,ATP是變構抑制劑,ADP是變構激活劑,Mg2+或K+可激活丙酮酸激酶的活性,胰島素可誘導PK的生成,烯醇式丙酮酸又可自動轉變成丙酮酸。
總結糖的無氧酵解 在細胞液階段的過程中,一個分子的葡萄糖或糖原中的一個葡萄糖單位,可氧化分解產生2個分子的丙酮酸,丙酮酸將進入線粒體繼續氧化分解,此過程中產生的兩對NADH+H+,由遞氫體α-磷酸甘油(肌肉和神經組織細胞)或蘋果酸(心肌或肝臟細胞)傳遞進入線粒體,再經線粒體內氧化呼吸鏈的傳遞,最后氫與氧結合生成水,在氫的傳遞過程釋放能量,其中一部分以ATP形式貯存。
在整個細胞液階段中的10或11步酶促反應中,在生理條件下有三步是不可逆的單向反應,催化這三步反應的酶活性較低,是整個糖的有氧氧化過程的關鍵酶,其活性大小,對糖的氧化分解速度起決定性作用,在此階段經底物水平磷酸化產生四個分子ATP。
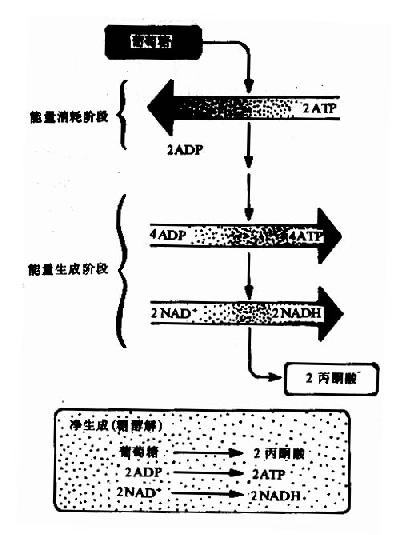
圖4-2 葡萄糖分解的兩個階段
總而言之,經過糖酵解途徑,一個分子葡萄糖可氧化分解產生2個分子丙酮酸。在此過程中,經底物水平磷酸化可產生4個分子ATP,如與第一階段葡萄糖磷酸化和磷酸果糖的磷酸化消耗二分子ATP相互抵消,每分子葡萄糖降解至丙酮酸凈產生2分子ATP,如從糖原開始,因開始階段僅消耗1分子ATP,所以每個葡萄糖單位可凈生成3分子ATP(圖4-2)。葡萄糖+2Pi+2NAD++2ADP→2丙酮酸+2ATP+2NADH+2H++2H2O
(三)丙酮酸在無氧條件下生成乳酸
氧供應不足時從糖酵解途徑生成的丙酮酸轉變為乳酸。缺氧時葡萄糖分解為乳酸稱為糖酵解(glycolysis),因它和酵母菌生醇發酵非常相似。丙酮酸轉變成乳酸由乳酸脫氫酶(lactatedehydrogenase)催化丙酮酸乳酸脫氫酶乳酸在這個反應中丙酮酸起了氫接受體的作用。由3-磷酸甘油醛脫氫酶反應生成的NADH+H+,缺氧時不能經電子傳遞鏈氧化。正是通過將丙酮酸還原成乳酸,使NADH轉變成NAD+,糖酵解才能繼續進行。
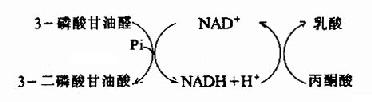
乳酸脫氫酶是由M和H二種亞基構成的四聚體,組成5種同工酶。這些同工酶在組織中分布不同,對丙酮酸的KM也有較大差異。H4主要分布在心肌。它的酶動力學參數表明H4有利于催化乳酸氧化成丙酮酸。所以心肌進行有氧氧化而且能利用乳酸作為燃料。骨骼肌中為M4型。它對反應方面無傾向性,但肌細胞內底物的濃度有利于生成乳酸。
(四)糖酵解及其生理意義
糖酵解是生物界普遍存在的供能途徑,但其釋放的能量不多,而且在一般生理情況下,大多數組織有足夠的氧以供有氧氧化之需,很少進行糖酵解,因此這一代謝途徑供能意義不大,但少數組織,如視網膜、睪丸、腎髓質和紅細胞等組織細胞,即使在有氧條件下,仍需從糖酵解獲得能量。
在某些情況下,糖酵解有特殊的生理意義。例如劇烈運動時,能量需求增加,糖分解加速,此時即使呼吸和循環加快以增加氧的供應量,仍不能滿足體內糖完全氧化所需要的能量,這時肌肉處于相對缺氧狀態,必須通過糖酵解過程,以補充所需的能量。在劇烈運動后,可見血中乳酸濃度成倍地升高,這是糖酵解加強的結果。又如人們從平原地區進入高原的初期,由于缺氧,組織細胞也往往通過增強糖酵解獲得能量。
在某些病理情況下,如嚴重貧血、大量失血、呼吸障礙、腫瘤組織等,組織細胞也需通過糖酵解來獲取能量。倘若糖酵解過度,可因乳酸產生過多,而導致酸中毒。
(五)糖酵解的調節
正常生理條件下,人體內的各種代謝受到嚴格而精確的調節,以滿足機體的需要,保持內環境的穩定。這種控制主要是通過調節酶的活性來實現的。在一個代謝過程中往往催化不可逆反應的酶限制代謝反應速度,這種酶稱為限速酶。糖酵解途徑中主要限速酶是己糖激酶(HK),磷酸果糖激酶-1(PFK-1)和丙酮酸激酶(PK)。
1.激素的調節
胰島素能誘導體內葡萄糖激酶、磷酸果糖激酶、丙酮酸激酶的合成,因而促進這些酶的活性,一般來說,這種促進作用比對限速酶的變構或修飾調節慢,但作用比較持久。
2.代謝物對限速酶的變構調節
上述三個限速酶中,起決定作用的是催化效率最低的酶PFK-1。其分子是一個四聚體形式,不僅具有對反應底物6-磷酸果糖和ATP的結合部位,而且尚有幾個與別位激活劑和抑制劑結合的部位,6-磷酸果糖、1,6二磷酸果糖、ADP和AMP是其激活劑,而ATP、檸檬酸等是其抑制劑,ATP既可作為反應底物又可作為抑制劑,其原因在于:此酶一個是與作為底物的ATP結合位點,另一個是與作為抑制劑的ATP結合位點,兩個位點對ATP的親和力不同,與底物的位點親和力高,抑制劑作用的位點親和力低。對ATP有兩種結合位點,這樣,當細胞內ATP不足時,ATP主要作為反應底物,保證酶促反應進行,而當細胞內ATP增多時,ATP作為抑制劑,降低了酶對6-磷酸果糖的親和力。
它在體內也是由6-磷酸果糖磷酸化而成,但磷酸化是在C2位而不是C4位,參與的酶也是另一個激酶,磷酸果糖激酶-2(PFK-2)。
2,6-二磷酸果糖可被二磷酸果糖磷酸酶-2去磷酸而生成6-磷酸果糖,失去其調節作用。2,6-二磷酸果糖的作用在于增強磷酸果糖激酶-1對6-磷酸果糖的親和力和取消ATP的抑制作用(圖4-3)。
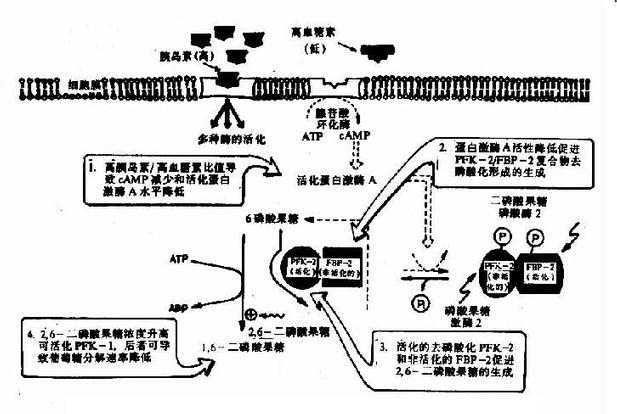
圖4-3 胰島素濃度升高對肝細胞內2,6-二磷酸果糖濃度的影響
臨床上丙酮酸激酶異常,可導致葡萄糖酵解障礙,紅細胞破壞出現溶血性貧血。
| 關于“生物化學與分子生物學/糖酵解途徑”的留言: | |
|
目前暫無留言 | |
| 添加留言 | |